
Статья 7
О ВЛИЯНИИ СПОСОБОВ ЗРИТЕЛЬНОГО
ОПОЗНАНИЯ
НА ФУНКЦИОНАЛЬНОЕ СОСТОЯНИЕ
ЧЕЛОВЕКА-ОПЕРАТОРА
Рубахин В. Ф., Кухтина Л. Г.
Стр. 51-59
Изучение перцептивных действий в
контексте инженерно-психологических исследований представляет большой интерес в
связи с тем, что существует целый ряд операторских профессий, для которых
заданное качество перцептивных действий является необходимым условием высокой
продуктивности деятельности.
При изучении зрительного опознания
основное внимание уделялось установлению его механизмов и способов их
функционирования, закономерностям опознания зрительных стимулов различной степени
сложности, выработке эффективных навыков опознания и т. д. [1, 2, 4, 8 и др.].
Значительно меньше разработаны вопросы, связанные с изучением влияния различных
способов и механизмов опознания на динамику функционального состояния (ФС)
человека-оператора. Как известно [6], высокая продуктивность деятельности может
быть достигнута в том случае, когда имеется соответствие условий деятельности
функциональным возможностям человека. Индикатором степени соответствия
указанных факторов является состояние напряженности, рассматриваемое как
ведущее ФС человека, сопровождающее любую целенаправленную деятельность и сопровождающееся
мобилизацией нервных и соматовегетативных функций организма [3]. В настоящее
время выделены две формы состояния напряженности: продуктивная и непродуктивная.
Критерием их выделения является степень мобилизации психофизиологических
функций организма [3, 7]. Количественной мерой дифференциации указанных форм
напряженности служит психофизиологическая цена деятельности (ПФ-цена) [6]. Как
было установлено [3, 6], продуктивная (оптимальная) форма напряженности может
быть достигнута в том случае, когда высокая эффективность деятельности
человека-оператора сопровождается ее низкой психофизиологической ценой. При
данной форме напряженности обеспечивается длительная и эффективная
работоспособность. Высокая ПФ-цена деятельности соответствует непродуктивным
(неоптимальным) формам напряженности, при которых нарушается соответствие
условий труда функциональным возможностям человека-оператора.
Таким образом, оптимизация
зрительной перцептивной деятельности человека-оператора (в частности, процессов
опознания) невозможна без установления факторов, влияющих на ее продуктивность.
Необходимость такого рода исследований связана также с тем, что в подавляющем
числе операторских профессий процесс опознания осуществляется в условиях
ограничения либо времени предъявления информации, либо возможности отвлечения
от основного вида деятельности (например, управления движущимся объектом).
Дефицит времени опознания предъявляет большие требования как к качеству
опознания, так и к степени мобилизации психофизиологических функций организма.
Для прогнозирования длительной продуктивной деятельности человека-оператора,
связанной с опознанием сложной по составу зрительной информации в условиях
дефицита времени, необходимо выяснить, какие способы и механизмы опознания
обусловливают низкую ПФ-цену деятельности, т. е. сопровождаются адекватной
мобилизацией соответствующих физиологических систем. Решение этого вопроса не
только позволит установить наиболее предпочтительные способы зрительного
опознания при различных временных режимах предъявления информации, но и даст возможность
использовать диагностику той или иной формы состояния напряженности для оценки
степени соответствия механизмов перцептивной деятельности условиям ее
реализации. Приведенное ниже экспериментальное исследование посвящено изучению
влияния механизмов опознания многомерных зрительных стимулов при различных
временных режимах предъявления информации на динамику состояния напряженности
человека-оператора.
Методический комплекс состоял из
сочетания метода тахистоскопического предъявления информации с заданными
периодами экспозиции и полиэффекторного метода регистрации психофизиологических
функций. Специфика изучаемой деятельности определила состав одновременно регистрируемых
психофизиологических параметров. При исследовании зрительного опознания такими
информативными параметрами являются электроокулограмма (специфический индикатор
ФС), кожно-гальваническая реакция (КГР) и электромиограмма (ЭКГ)
артикуляционных мышц (неспецифические индикаторы ФС) [9]. Тестовый материал
представлял собой модификацию часто применяемых при такого рода исследованиях
черно-белых таблиц и состоял из 72 матриц. Каждая матрица содержала 16 клеток
(4Х4), заполненных согласными буквами русского ·алфавита. Стимул, подлежащий
опознанию, имел следующие признаки:
1. Буквенный состав. Стимульную
совокупность составляли четыре буквы (ПВХН и ТБЦМ).
2. Конфигурация. Для обоих наборов
буквы, составляющие стимул, располагались в постоянно заданном порядке - три
буквы последовательно и одна сбоку. Таким образом, конфигурация стимула
напоминала ход шахматного коня. Имелось восемь возможных вариантов пространственного
расположения каждой стимульной совокупности на клетках матрицы (четыре
горизонтальных и четыре вертикальных). Имелось три условия предъявления:
«положительный стимул» (S+); «отрицательный стимул» (S_), отличавшийся тем, что буквы, составляющие стимульную
совокупность, были расположены либо в строку, либо столбцом, либо по диагонали
клеток матрицы; «шум» (Ш), когда буквы стимула располагались по клеткам в
случайном порядке. Тестовый материал предъявлялся в случайном порядке.
Исследование проводилось на экспериментальном стенде с автоматизированной
регистрацией времени решения задач, дополненном психофизиологической
аппаратурой. Испытуемый находился в звукоизолированной камере. Работа
проводилась в два этапа. На первом этапе оценивалась только эффективность опознания
тестовых объектов (по показателям: скорости и точности). На втором этапе
проводилась комплексная объективная оценка восприятия тестовых объектов с
помощью электрофизиологических показателей с целью определения состояния
зрительной системы и функционального состояния в целом. Процедура эксперимента
состояла в следующем. Испытуемому на экране предъявлялось тестовое изображение
с временами экспозиции (tэ), постоянными для каждой
экспериментальной серии. Времена экспозиции составляли 300, 500, 1000, 1500 и 3000
мс. До и после изображения предъявлялась маскировочная матрица, заполненная
буквой А. Испытуемый находился на расстоянии 700 мм от экрана. Размер
изображения составлял 80. При опознании предъявленного изображения
испытуемый нажимал кнопку микропереключателя (для остановки миллисекундомера) и
заполнял формуляр, указывая в нем конфигурацию и расположение стимула, либо
«шум». Формуляр представлял собой пустые матрицы. Для каждого времени экспозиции
с испытуемыми проводилось по две подготовительные и три основные
экспериментальные серии. На втором этапе процедура предъявления тестового
материала оставалась прежней, однако регистрация глазодвигательной активности
(ГДА), КГР, ЭМГ артикуляционных мышц приводила к изменению подготовительного
этапа, так как испытуемому необходимо было адаптироваться к условиям
электрофизиологического исследования. Анализировались следующие данные
эксперимента:
- точность опознания (процент
правильных ответов);
- скорость опознания (время от
начала предъявления до моторного ответа);
- характер ошибок опознания;
- показатели ГДА (общее время,
среднее количество скачков глаз, среднее значение амплитуды скачков, средняя длительность
фиксаций);
- характеристики КГР (латентный
период, амплитуда и длительность реакций, количество фаз КГР, количество
реакций);
- характеристики ЭМГ
артикуляционных мышц (количество и длительность фазических реакций, амплитуда
биоэлектрической активности артикуляционных мышц).
Анализ полученных
экспериментальных данных позволил выделить три труппы испытуемых, у которых
существенно различались как показатели эффективности опознания при различных
временах экспозиции, так и показатели, характеризующие функциональное
состояние.
Данные
эффективности опознания тестового материала по выделенным группам приведены на
рис, 1. На рис. 2 приведены данные зависимости времени опознания положительного
и отрицательного стимула и шума от времени экспозиции. Как видно из приведенных
данных, первая группа испытуемых характеризуется наибольшими значениями
показателей скорости и точности опознания при всех временах экспозиции стимула.
Соотношение времени опознания стимулов и шума (Ш) было следующим: ВО (S_) ≤ BO
(S+) < ВО (Ш), Время опознания стимулов в диапазоне tэ от 1000 от
3000 мс было меньше времени экспозиции (рис. 2, а). Вторая группа испытуемых
характеризовалась более низкими значениями скорости и точности опознания
тестового материала. Лишь при tэ 3000 мс время опознания стимулов
было меньше заданного времени экспозиции. Соотношение времен опознания стимулов
и шума было таким же, как у испытуемых первой группы, однако время опознания
отрицательного стимула становилось меньше времени опознания положительного
только при tэ 3000 мс (рис. 2, б). Самые низкие показатели скорости
и точности опознания при всех tэ наблюдались у испытуемых третьей
группы. Время опознания тестового материала во всех случаях превышало время
экспозиции. Соотношение времен опознания положительного и отрицательного стимула
и шума можно выразить следующим образом: ВО (S_) = ВО (S+) ≤ BO(Ш) (рис. 2, в).
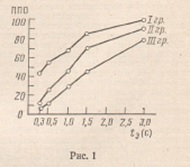
Рис. 1. Зависимость точности опознания от длительности экспозиции стимула
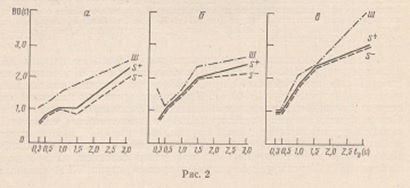
Рис. 2. Зависимость времени опознания от длительности экспозиции стимула. а - первая, б - вторая, в - третья группа испытуемых
Анализ результатов точности опознания каждого из восьми вариантов стимула показал, что испытуемые первой группы практически с одинаковой точностью опознавали все варианты при tэот 500 до 3000 мс. У испытуемых второй и третьей групп зависимость эффективности опознания вариантов стимула от времени экспозиции была выражена в большей мере, особенно при tэ ниже 1500 мс. Так, разница в точности опознания между 1, 2, 5, 7-м и 3, 6, 8-м вариантами достигала 30-40%. Место расположения стимула на клетках матрицы также в большей мере оказывало влияние на эффективность опознания испытуемых второй и третьей групп. Предъявление стимула, расположенного на нижних клетках матрицы, было более замедленным (время опознания увеличивалось в среднем на 800 мс) и менее точным (процент правильных ответов уменьшался на 40 %), чем при опознании аналогичных стимулов, расположенных на верхних клетках матрицы и в ее центре.
При рассмотрении
ошибок опознания было установлено, что испытуемые первой группы допускали в
основном ошибки типа «сдвиг стимула по клеткам матрицы» без нарушения его пространственной
ориентации. Испытуемые второй группы вместе с этими ошибками неправильно
определяли пространственную ориентацию стимула, поворачивая его зеркально по
отношению к предъявленному. Испытуемые третьей группы указывали «зону
присутствия стимула» с частичным заполнением клеток матрицы буквами стимула.
Наряду с этим уменьшение времени экспозиции стимула в большей, мере приводило к
появлению ошибок в опознании пространственной ориентации стимула у испытуемых
третьей группы, испытуемые первой группы допускали такого рода ошибки только
при tэ ниже 1000 мс (рис. 3).
Результаты регистрации
глазодвигательной активности наряду с данными эффективности позволили
установить, каким способом испытуемые той или иной группы опознавали
предъявленный материал, так как характер ГДА у них оказался различным. Из
данных, приведенных в табл. 1, видно, что у испытуемых первой группы при tэот 500 до 1500 мс зарегистрирован самый короткий период ГДА, наименьшее
количество скачков глаз и наименьшая их амплитуда. Максимальный период ГДА,
большое количество скачков глаз и максимальная их амплитуда зарегистрированы
при указанных временах экспозиции у испытуемых третьей группы. Анализ ГДА
показал, что при tэ 1500 и 3000 мс все испытуемые опознавали стимул
за счет последовательной смены точек фиксации на экспозиционном поле. При tэниже 1500 мс испытуемые первой группы опознают стимул в среднем за две
фиксации, а соотношение времен опознания положительного и отрицательного
стимулов позволяет предположить, что стимулы опознаются симультанно, т. е. все
признаки стимула обрабатываются одновременно. Такой же вывод можно сделать и
относительно испытуемых второй группы, однако данные эффективности опознания,
характеризующие качество их работы, свидетельствуют о недостаточной
сформированности у них процесса параллельного сличения признаков стимула с
эталоном. Испытуемые третьей группы последовательно опознавали все элементы
стимула, о чем свидетельствует большое число скачков глаз и минимальная
эффективность опознания особенно на коротких временах экспозиции. Таким
образом, при малых временах экспозиции стимула (от 300 до 1500 мс) наиболее
эффективным оказался симультанный способ опознания признаков стимула и менее
эффективным - последовательный (сукцессивный).
Оценка функционального состояния
испытуемых по данным КГР показала следующее. Наибольшая амплитуда реакции,
максимальная длительность и минимальный латентный период реакции наблюдались у
всех испытуемых при tэ 500 мс. Увеличение времени экспозиции привело
к различным изменениям в поведении указанных характеристик. Самые низкие
значения амплитуды (А) и длительности реакции (Т) оказались у испытуемых первой
группы, причем КГР в этой группе наблюдались лишь в 75 % случаев, особенно мало
их было при tэ 3000 мс. Испытуемым данной группы были присущи в
основном одно- и двух- фазные КГР. У испытуемых второй и третьей групп КГР
оказалась практически неугасимой: во второй группе были
зарегистрированы двух - четырехфазные реакции, в третьей - многофазные. Если у
испытуемых первой и второй групп: КГР возникали в постэкспозиционном периоде, то
у испытуемых третьей группы КГР следовали непосредственно за появлением стимула
(при tэ 500 и 1000 мс) или часто первая волна реакции совпадала с
началом экспозиции стимула. У испытуемых третьей группы наблюдалось также
появление первой волны КГР с латентным периодом, не превышавшим 1 с от момента
предъявления стимула (табл. 2).
Таблица
1
Показатели ГДА по
трем группам испытуемых
|
Группа
испытуемых
|
500 мс |
1000 мс |
||||||
|
Т, мс |
Nск |
tф, мс |
А, мкв |
Т, мс |
Nск |
tф, мс |
А, мкв |
|
|
I
II
III |
850 780 950
1100 1100 1100
1100 1100 1100
|
2,5 2,2 3,0
3,2 3,2 3,2
3,2 3,2 3,2 |
280 280 300
300 300 300
300 300 300 |
42,0 42,0 46,0
67,0 57,0 57,0
66,0 66,0 66,0 |
1150 1000 1350
1450 1450 1620
1900 2000 2100
|
3,1 2,5 3,5
4,0 4,0 5,1
5,9 6,7 6,9 |
300 300 300
320 320 310
300 300 300
|
42,0 43,0 50,0
60,0 60,0 60,0
70,0 71,0 71,0
|
|
Группа
испытуемых
|
1500 мс |
3000 мс |
||||||
|
Т, мс |
Nск |
tф, мс |
А, мкв |
Т, мс |
Nск |
tф, мс |
А, мкв |
|
|
I
II
III
|
1650 1550 2050
2000 2000 2450
2500 2550 2550 |
5,6 4,6 6,0
4,8 4,8 6,8
6,8 6,8 6,8 |
280 250 300
380 380 350
350 360 360 |
56,0 55,0 60,0
63,0 63,0 70,0
70,0 70,0 70,0 |
2400 2250 2850
2450 2450 2780
3000 3000 3800 |
7,0 6,5 9,4
7,8 7,8 8,2
9,2 9,2 11,2 |
300 300 300
300 300 300
300 300 320 |
61,0 61,0 64,0
60,0 60,0 62,0
62,0 62,0 66,0 |
Быстрое угасание КГР при tэот 3000 до 1000 мс зарегистрировано у испытуемых первой группы. Об этом
свидетельствовало исчезновение высокоамплитудных КГР и увеличение длительности
реакции. В то же время при tэ 500 и 1000 мс имевшиеся определенные
трудности в опознании, обусловленные кратковременным предъявлением стимула,
требовали поддержания необходимого функционального тонуса, способствующего
эффективному осуществлению процесса опознания. В связи с этим при указанных
временах экспозиции КГР оказалась достаточно выраженной, хотя носила тонический
характер, так как возникала только в постэкспозиционном периоде. Наряду с этим
не было выявлено корреляции между характеристиками КГР, предъявленным вариантом
стимула и местом его расположения в матрице.
У испытуемых второй группы
латентный период КГР был меньше, чем у испытуемых первой группы. Имела место
значительная длительность двухфазных реакций при высоких значениях амплитуд.
Наблюдалась выраженная зависимость характеристик КГР от времени экспозиции, что
свидетельствует о связи данной реакции не только с поддержанием функционального
тонуса, но и со сложностью опознания стимула при кратковременных экспозициях.
Следует отметить также тот факт, что даже при tэ 3000 мс значения
латентного периода, длительности и амплитуды КГР у испытуемых второй группы
превышают соответствующие значения испытуемых первой группы, зарегистрированные
при tэ 500 мс. У испытуемых второй группы наблюдалась определенная
зависимость таких характеристик КГР, как латентный период и амплитуда реакции
от варианта предъявленного стимула. При предъявлении «легких» вариантов (1, 2,
5 и 7) латентный период реакции у всех испытуемых был больше (на 30%) и
амплитуда меньше (на 40%), чем при предъявлении «сложных» вариантов (3 и 8).
При предъявлении стимула в центре и в верхней части матрицы амплитуда реакции была
на 20% ниже, чем при расположении стимула в нижней части матрицы, при этом
длительность латентного периода возрастала в среднем на 20 %. Это дает возможность
предположить, что повышенный уровень состояния напряженности, наблюдаемый у
испытуемых второй группы, связан как с временными режимами предъявления информации,
так и со сложностью процесса ее опознания.
Таблица
2
Изменение основных характеристик
КГР
в зависимости от
времени экспозиции
|
tэ ,
мс |
Регистрируемые параметры |
Группы испытуемых |
||
|
I |
II |
III |
||
|
500
1000
1500
3000 |
Т,
с ЛВ,
с А,
мкв Количество реакций, %
Т,
с ЛВ,
с А,
мкв Количество реакций, %
Т,
с ЛВ,
с А,
мкв Количество реакций, %
Т,
с ЛВ,
с А,
мкв Количество реакций, % |
3,0 3,0 600 75
3,5 3,0 550 70
5,0 4,5 400 55
4,5 5,0 400 47 |
9,0 2,0 1200 95
7,0 3,0 1010 95
6,6 2,5 900 75
6,0 4,2 800 94 |
10,0 1,0 2000 100
8,0 1,0 1880 100
9,5 1,0 1600 100
8,0 1,0 1020 100 |
В третьей группе испытуемых наблюдались
КГР максимальной выраженности, причем они носили в основном физический
характер, так как возникали в период экспозиции стимула (при tэ1500, 3000 мс) или непосредственно после его окончания. КГР отличались высокой
амплитудой, малой длительностью каждой фазы и большим временем угасания
реакции. Максимальная амплитуда КГР наблюдалась при предъявлении 3, 4-го и 8-го
вариантов стимула и отличалась от таковой при предъявлении 1, 2-го и 5-го
вариантов в среднем на 35%. Значения амплитуды КГР при предъявлении стимула в
центре матрицы были минимальными и отличались от соответствующих значений при
расположении стимула в правом верхнем и нижних секторах матрицы в среднем на
30%. Изменений значений латентного периода КГР у третьей группы испытуемых не
наблюдалось. Анализ характеристик КГР у данной группы испытуемых показал, что в
этом случае она была связана, как со сложностью самого процесса опознания, так
и с повышенным уровнем эмоциональной напряженности, обусловленным низкой
эффективностью деятельности практически при всех вpeмeнax экспозиции стимула. Все
изложенное позволяет предположить следующее. Изменения со стороны вегетативной
нервной системы у испытуемых первой группы были связаны в основном с
поддержанием того функционального тонуса, который обеспечивал эффективное
опознание при малых временах экспозиции стимула; они свидетельствуют о
состоянии адекватной (продуктивной) напряженности при опознании стимулов в
диапазоне времен экспозиции от 1000 до 3000 мс. Изменения, наблюдаемые у второй
и третьей групп испытуемых, связаны с возрастанием сложности опознания при
малых временах экспозиции стимула и свидетельствуют о состоянии неадекватной
(непродуктивной) напряженности при tэ ниже 3000 мс. В большей
степени данное состояние выражено у испытуемых третьей группы.
Таблица
3
Данные ЭМГ
артикуляционных мышц
|
Время экспозиции, мс |
Группы испы-туемых |
Количество фазических реакций, % |
Латентный период реакции, с |
Средняя длительность реакции, с |
Средние значения Ам , мкв |
||
|
фон |
1* |
2** |
|||||
|
3000
1500
1000
500 |
I II III
I II III
I II III
I II III
|
0 0 80
0 40 100
0 65 100
0 87 100
|
0 0 2,8
0 2,9 3,7
0 1,6 2,0
0 1,2 1,9
|
0 0 4,3
0 4,7 5,8
0 5,2 5,7
0 8,2 7,0 |
20 20 15
20 20 15
20 20 15
20 20 15 |
40 30 150
45 35 80
45 40 100
50 40 100 |
40 30 300
45 80 220
45 100 300
50 190 300 |
*Период от начала экспозицин до моторного
ответа
**Период
после моторного ответа (заполнение формуляра).
Как указывалось [3,
5, 9], по данным ЭМГ артикуляционных мышц можно судить как о наличии
затруднений, появляющихся в процессе опознания, так и об уровне функционального
состояния напряженности, возникающем в процессе данной деятельности.
Особый интерес представляла
регистрация биоэлектрической активности (БА) артикуляционных мышц в связи с
тем, что испытуемые не давали ответ в речевой форме. Поэтому наличие скрытой
речевой активности как в период опознания стимула, так и в период заполнения
формуляра позволяло констатировать определенные затруднения, возникавшие в
процессе опознания стимулов при различных временах экспозиции. Данные суммарной
БА артикуляционных мышц, зарегистрированные в процессе эксперимента,
сравнивались с фоновыми значениями, что давало возможность установить общий
уровень состояния напряженности при различных временных режимах предъявления
информации. Анализ характеристик ЭМГ позволил выделить наиболее информативные
из них в отношении исследуемой деятельности. Такими характеристиками оказались:
средняя максимальная амплитуда ЭМГ (Ам), латентный период
возникновения скрытой речевой активности и ее длительность. БА круговой мышцы
рта имела в большинстве случаев тонический характер. Однако наблюдались
реакции, характеризующиеся достаточно выраженной фазической составляющей. Для
каждого времени экспозиции стимула подсчитывалось среднее количество
скрыторечевых реакций (в процентах по отношению к общему числу предъявленных
стимулов). Данные регистрации ЭМГ артикуляционных мышц при различных временах
экспозиции приведены в табл. 3. Они свидетельствуют о различном характере БА,
связанном как со способом опознания стимулов, так и с влиянием временных
режимов предъявления информации. У испытуемых первой группы скрыторечевые
реакции не наблюдались ни при одном из времен экспозиции. Значения Амв процессе эксперимента превышали аналогичные фоновые значения в 2-2,5
раза. Причем общая тенденция заключалась в следующем: чем меньше время
экспозиции, тем больше значение Ам что, на наш взгляд, связано с
поддержанием оптимального функционального тонуса, способствующего эффективиой
зрительной деятельности, протекающей в режиме дефицита времени. У всех
испытуемых первой группы наблюдалась относительно стабильная картина БА
артикуляционных мышц в процессе эксперимента с некоторым ее повышением в конце
каждого опыта, что, по-видимому, связано с компенсацией возникающего утомления:
Отсутствие скрыторечевых реакций у испытуемых данной группы, относительная
стабильность уровня БА и небольшой диапазон изменений значений Ампри различных временах экспозиции могут свидетельствовать как о высокой степени
автоматизации перцептивного навыка, так и о том, что опознавательные действия
совершаются в образном плане. У испытуемых второй группы картина изменений БА
артикуляционных мышц несколько иная. При tэ 3000 мс у испытуемых этой
группы скрыторечевых реакций не наблюдалось. Однако было выявлено некоторое
увеличение значений Ам в период после моторного ответа по сравнению
с аналогичными значениями, зарегистрированными в период опознания. При tэ1500 мс в 40% случаев были зарегистрированы скрыторечевые реакции, возникающие в
период заполнения формуляра. Количество подобных реакций возрастало с уменьшением
времени экспозиции стимула: при t. 1000 мс их было 65%, при tэ 500
мс-87%. Средняя максимальная амплитуда ЭМГ также возрастала с уменьшением
времени экспозиции при выраженной тенденции увеличения длительности
скрыторечевых реакций. Значения Ам в период опознания стимула
незначительно отличались от фоновых (в среднем в 1,5-2 раза). Появление
скрыторечевых реакций у данной группы испытуемых было связано как со степенью
сложности опознания отдельных вариантов стимула, так и с определением места его
расположения в матрице. При tэ 1500 мс скрыторечевая активность
наблюдалась при предъявлении «сложных» вариантов стимула (6 и 8) на нижних
клетках матрицы. При tэ 1000 и 500 мс скрыторечевые реакции
возникали при предъявлении 3, 4, 6-го и 8-го вариантов стимула в правом верхнем
и нижних секторах матрицы. При tэ от 1500 до 500 мс скрыторечевые
реакции появлялись как в случаях предъявления матриц, содержащих стимул, так и
в случаях предъявления «пустых» матриц. при всех tэ скрыторечевая
активность наблюдалась у испытуемых данной группы только в период после
моторного ответа, что, по-видимому, связано с процессом принятия решения о
наличии или отсутствии стимула, указанием его конфигурации и места расположения
в матрице. При этом в большинстве случаев возникновение скрыторечевых реакций
сопровождалось появлением КГР. Так, при tэ 1500 мс 60% скрыторечевых
реакций сопровождалось КГР, при tэ 1000 мс такие совпадения
наблюдались в 80% случаев, при tэ 500 мс - в 98% случаев. Таким
образом, появление скрыторечевой активности у испытуемых второй группы,
опознававших стимул путем параллельной обработки его признаков (симультанно),
зависело как от временных режимов экспозиции тест-объектов, так и от
характеристик самого стимула и места его расположения в матрице, что
свидетельствует о недостаточной степени сформированности данного способа
опознания в отличие от испытуемых первой группы.
У испытуемых третьей группы
скрыторечевая активность наблюдалась при всех временах экспозиции тестового
материала. Причем эти реакции были зарегистрированы как в период опознания
стимула (до моторного ответа), так и в период заполнения формуляра. При tэ3000 мс БА артикуляционных мышц фазического характера наблюдалась в 26%
случаев в период опознания и в 30.% случаев в период заполнения формуляра. При
этом средние значения Ам, ЭМГ в период заполнения формуляров были
вдвое выше, чем в период опознания. Эта тенденция сохранилась и при остальных
временах экспозиции, причем чем оно было меньше, тем на большую величину
различались данные средние значения Ам. При t. 1500, 1000 и 500 мс
скрыторечевая активность наблюдалась в 100 % случаев и появлялась после
моторного ответа. С уменьшением времени экспозиции латентный период реакции сокращался,
а ее длительность возрастала. Наличие скрыторечевой активности в период
экспозиции 3000 мс было характерно только для третьей группы испытуемых. Таким
образом, последовательный способ опознания признаков стимула, характерный для
данной группы испытуемых, сопровождался ярко выраженной скрыторечевой
активностью, особенно при малых временах экспозиции. У испытуемых третьей
группы также наблюдалось возникновение КГР, совпадающих по времени с повышением
БА артикуляционных мышц. При tэ 3000 мс этот факт был
зарегистрирован в 78% случаев, при tэ 1500 мс - в 80.% случаев и при
tэ 1000 и 500 мс - в 100 % случаев. Таким образом, в результате проведенного
исследования установлено следующее.
Каждый из выявленных способов
опознания сложных зрительных стимулов при различных временных режимах их
предъявления характеризуется наряду с показателями эффективности деятельности
различными показателями функционального состояния напряженности, т. е. различной
психофизиологической ценой деятельности. Так, в условиях кратковременного
предъявления информации наиболее эффективным является симультанный способ
опознания, при котором высокая результативность сопровождается умеренной
(адекватной) активацией функциональных физиологических систем, т. е. низкой
ПФ-ценой деятельности, что обусловливает возникновение состояния продуктивной
напряженности. Опознание посредством последовательного перебора признаков
стимула (сукцессивное) менее эффективно при малых временах экспозиции. Этот
способ характеризуется соотношением низкой результативности опознания с высоким
(неадекватным) уровнем активации функциональных физиологических систем, что
приводит к возникновению состояния непродуктивной напряженности. Высокая
степень автоматизации зрительного перцептивного навыка симультанного опознания способствует
снижению ПФ-цены деятельности и возникновению состояния продуктивной
напряженности. Исходя из изложенного, при оптимизации зрительной перцептивной
деятельности человека-оператора по опознанию сложной визуальной информации в
условиях дефицита времени целесообразны наиболее эффективные способы опознания,
способствующие длительной продуктивной работе при минимальных «психических» и
«органических» затратах на ее осуществление. Критерием для отбора в данном
случае могут служить показатели, характеризующие психофизиологическую цену
деятельности в сочетании с данными эффективности.
ЛИТЕРАТУРА
1.
Гunnеnрейтер Ю. Б. Движения человеческого глаза. М., 1978.
2.
Зuнченко В. П. Продуктивное восприятие. - Вопр. психологии, 1971, № 6.
3.
Методы и критерии оценки функционального комфорта. М., 1978 (Методические
материалы ВНИИТЭ).
4.
Подольскuй А. И. Формирование симультанного опознания. М., 1978.
5.
Соколов А. Н. Внутренняя речь и мышление. М., 1963.
6.
Чайнова Л. Д. Функциональный комфорт. Компоненты. Условия формирования.- Техническая
эстетика, 1983, № 1
7.
Чайнова Л. Д., Левшuнова Ж. В., Кашuрuна Л. В. О важности дифференцированной
оценки состояния напряженности. - В кн.: Проблемы функционального комфорта. М.,
1977.
8.
Шехтер М. С. Зрительное опознание. М., 1981.
Электронные журналы Института психологии РАН
Международная юбилейная научная конференция
7 - 8 декабря 2023 г., Москва
Сборник конференции молодых ученых, 2019
Примите участие в исследовании:

Моя экономическая жизнь в условиях пандемии COVID-19"
и поделитесь ссылкой на него с другими!
Ситуация пандемии COVID-19 - уникальна, требует изучения и осознания. Сроки для этого сжаты
Коллективная память о событиях отечественной истории

Психология глобальных рисков
